BMC Medicine volume 20, Article number: 494 (2022) Cite this article
Abstract
Obesity usually is accompanied by inflammation of abdominous tissue, with a salient relation of visceral fat. Chronic inflammation successful obese abdominous insubstantial is of a little people than acute immune activation for clearing the insubstantial from an infectious agent. It is the nonaccomplishment of adipocyte metabolic homeostasis that causes activation of nonmigratory immune cells for supporting insubstantial functions and regaining homeostasis. Initially, the excess influx of lipids and glucose successful the discourse of overnutrition is met by adipocyte maturation and proliferation. Eventual lipid overload of hypertrophic adipocytes leads to endoplasmic reticulum accent and the secretion of a assortment of signals causing accrued sympathetic tone, lipolysis by adipocytes, lipid uptake by macrophages, matrix remodeling, angiogenesis, and immune compartment activation. Pro-inflammatory signaling of adipocytes causes the nonmigratory immune strategy to merchandise accrued amounts of pro-inflammatory and different mediators resulting successful enhanced tissue-protective responses. With chronic overnutrition, these protective actions are insufficient, and decease of adipocytes arsenic good arsenic senescence of respective insubstantial compartment types is seen. This structural harm causes the look oregon merchandise of immunostimulatory compartment components resulting successful influx and activation of monocytes and galore different immune compartment types, with a publication of stromal cells. Matrix remodeling and angiogenesis is further intensified arsenic good arsenic perchance detrimental fibrosis. The accumulation of senescent cells besides whitethorn beryllium detrimental via eventual dispersed of senescence authorities from affected to neighboring cells by the merchandise of microRNA-containing vesicles. Obese visceral abdominous inflammation tin beryllium viewed arsenic an initially protective effect successful bid to header with excess ambient nutrients and reconstruct insubstantial homeostasis but whitethorn lend to insubstantial harm astatine a aboriginal stage.
Background
Fat insubstantial is interspersed with nonmigratory immune cells arsenic are each different coagulated tissues of the body. Activation of specified immune cells usually is accompanied by section oregon systemic inflammation of varying intensity. A cardinal diagnostic of inflammation is the accrued accumulation of cytokines, chemokines, and different immune mediators which hindrance to cognate receptors contiguous on/in galore compartment types successful the section insubstantial and passim the body. In summation to the relation of nonmigratory immune cells, astir each different compartment types of a insubstantial tin beryllium activated to merchandise pro-inflammatory mediators, usually astatine little levels that seen for “professional” immune cells. Mild responses see section insulin resistance, oxidative accent and altered compartment metabolism. Higher degrees of inflammation are characterized by the activation and infiltration of circulating immune cells which whitethorn origin section pain, edema, oregon fever [1, 2].
Triggers of nonmigratory immune compartment activation not lone comprise microbial oregon different infections but besides “sterile” stimuli specified arsenic metabolic, physical, oregon toxic accent starring to excess accumulation of oxygen radicals, reactive lipids, macromolecule aggregates, accent proteins oregon compartment necrosis followed by the merchandise of divers immunostimulatory compounds termed damage-associated molecular patterns [1].
The contiguous reappraisal describes changes successful visceral abdominous insubstantial successful effect to chronic overnutrition, the signals and compartment types progressive successful the aboriginal stages of insubstantial inflammation, and the progression to full-blown inflammation characterized by insubstantial harm and infiltration of circulating immune cells.
Main text
Resident immune cells successful thin visceral abdominous tissue
In visceral fat, members of the innate arsenic good arsenic adaptive immune strategy person been identified. These see macrophages, dendritic cells, granulocytes, innate lymphoid cells (ILCs and earthy slayer (NK) cells), and besides T and B cells [3] (Fig. 1). Single-cell transcriptome investigation of rodent thin visceral adipose insubstantial leukocytes identified 15 chiseled subpopulations [4]. In mean non-inflamed tissue, these cells are not lone immune guardians against corruption but besides enactment due insubstantial function. Many of these findings originate from studies successful experimental models, but wherever analyzed, akin physiological functions of immune cells person besides been reported successful humans [5]. For instance, macrophages grounds functional heterogeneity which includes the removal of dormant oregon apoptotic abdominous cells, remodeling the extracellular matrix and promoting angiogenesis [5]. A subtype of macrophages supports the power of lipid metabolism by uptake and digestion of lipids [6, 7]. Furthermore, the secretion of IL-27 appears to beryllium a large pathway of promoting thermogenesis successful abdominous cells [8]. Macrophages lend to the regularisation of thermogenesis successful effect to acold vulnerability [9]. There is nary homogeneous organisation of macrophage subtypes. For instance, successful quality subcutaneous tissue, spatial mapping identified macrophages with a M1-like phenotype associated with niches of adipocyte progenitor cells portion macrophages with a non-inflammatory phenotype were dispersed passim the abdominous insubstantial [10].

Network and physiological functions of nonmigratory immune cells successful thin visceral adipose tissue. In the lack of metabolic oregon inflammatory accent nonmigratory immune cells interact among themselves and with adipocytes and stromal cells to support due insubstantial functions. There are nary signature cytokines defining the attraction authorities of nonmigratory immune cells. Rather, the conception of a buffered strategy applies, without polarization towards a Th1/M1- oregon Th2/M2-like signifier oregon towards different biased authorities of immune reactivity. Cytokines, chemokines, acute signifier proteins, and different immune mediators are released successful tiny amounts mostly from nonmigratory immune cells but besides from mesenchymal stromal cells and adipocytes. Several macrophage subtypes beforehand matrix remodeling and angiogenesis, phagocytose dormant compartment and lipid aggregates, and beforehand adipocyte thermogenesis. ILC2 besides supports adipocyte thermogenesis and stimulates physiological eosinophil functions. Regulatory T cells beforehand insubstantial repair and interact with macrophages and different immune compartment types to support a non-inflammatory state. Low-level secretion of immune mediators by macrophages, dendritic cells, and different immune compartment types specified arsenic ILC2s, iNKTs, Th2 cells, γδT cells, B-1b cells, and eosinophils helps to forestall immune compartment activation. For amended readability, lone a fewer cardinal intercellular signals are included successful the scheme. ATM, adipose insubstantial macrophage; DC, dendritic cell; IL, interleukin; ILC, innate lymphoid cell; iNKT, innate earthy slayer T cell; MetEnk, methionine-enkephalin peptides; NK, earthy slayer cell
ILC2 cells lend to the regularisation of vigor expenditure by promoting the differentiation of beige adipocytes from adipocyte precursors oregon beiging of achromatic abdominous cells successful visceral tissues via upregulation of uncoupling macromolecule 1 (UCP1 by enkephalin peptides [11, 12]. ILC2-derived IL-13 helps to forestall pro-inflammatory activation of macrophages, dendritic cells, ILC1, and earthy slayer cells. By secreting IL-5, ILC2 cells beforehand anti-inflammatory eosinophil activity.
Conventional dendritic cells grounds a tolerogenic phenotype, characterized by IL-10 accumulation and suppression of Th1-promoting enactment by upregulated look of peroxisome proliferator-activated receptor gamma (PPAR-γ) [13]. Regulatory T cells (Treg) correspond the large CD4-positive T compartment type, they enactment successful insubstantial repair and sphere Glut-4 look by adipocytes [14, 15]. Interestingly, the bulk of Tregs look to beryllium oligoclonal successful mice arsenic indicated by chiseled T compartment receptor repertoires. There whitethorn beryllium MHC II-dependent antigen designation involved, arsenic suggested by the adjacent relation with nonmigratory macrophages and dendritic cells [16]. The superior relation of Tregs astir apt is to support different immune compartment types successful a neutral physiological state, i.e., preventing immune activation either towards a pro-inflammatory oregon a Th2-like state.
Resident B-1b lymphocytes secrete earthy IgM antibodies and beforehand adipose physiological functions by suppressing B-2 cells, successful mice and humans [17]. In addition, B-1 cells comprise the large compartment benignant of fat-associated lymphoid clusters which look to lend to humoral immune responses to peritoneal antigens [18]. Lymphoid clusters successful mice and humans are besides a affluent root of Th2-like cytokines released from innate Th2-like lymphoid cells [19, 20]. Fat-associated lymphoid clusters specified arsenic milky spots connected the omentum aboveground astir apt service immune functions of the peritoneal cavity alternatively than supporting physiological abdominous insubstantial functions. Indeed, the numbers of milky spots summation during peritoneal inflammation successful effect to section TNFα and innate earthy slayer T compartment enactment [20, 21]. Studies successful mice suggest that sympathetic innervation is promoted by γδT cells by signaling via the IL-17 receptor C to induce TGFß1 accumulation by parenchymal cells [22]. Further, sympathetic neuron-associated macrophages (SAMs) modulate neuron maturation and modulate adrenergic signaling [23].
Based mostly connected carnal studies, the continuous merchandise of anti-inflammatory mediators from macrophages, dendritic cells, Th2-cells, γδT cells, eosinophils, mucosa-associated invariant T cells (MAIT), and invariant earthy slayer T cells appears to further assistance support metabolic homeostasis [14, 24,25,26,27,28,29,30,31,32] (Fig. 1). The enactment of insubstantial functions by nonmigratory immune cells involves interactions with non-immune insubstantial cells including adipocytes, endothelial cells, neurons, fibroblasts, and different mesenchymal stromal cells [3, 33, 34].
In the lack of immunologic stimuli, immune mediator secretion from nonmigratory immune cells and different abdominous insubstantial cells is low. The section immune milieu is good buffered, i.e., determination is neither a dominance of Th1/M1-like nor Th2/M2-like immune reactivity. In this context, it is important to enactment that determination is interdependence of Th1/M1- and Th2/M2-associated cytokines. For instance, the Th1-type cytokine TNFα stimulates the accumulation of the Th2/Treg-associated cytokine IL-10 which successful crook downregulates TNFα. Further, pro-inflammatory TNFα and IL-17A induce counterregulatory IL-33 for the stimulation of anti-inflammatory Tregs and ILC2s [15, 35, 36].
Taken together, successful thin visceral adipose tissue, determination is simply a physiological web of adipocytes, stromal cells, and immune cells. The nonmigratory immune strategy is not dormant but supports wide insubstantial functions. There are nary signature cytokines defining the attraction authorities of nonmigratory immune cells. Rather, the conception of a buffered strategy applies, without polarization towards a Th1/M1- oregon Th2/M2-like signifier oregon towards different biased authorities of immune reactivity. Cytokines, chemokines, acute signifier proteins, and different immune mediators are released successful tiny amounts mostly from nonmigratory immune cells but besides from mesenchymal stromal cells and adipocytes [37, 38].
From thin to obese visceral abdominous tissue
The superior origin of progression from thin to obese visceral abdominous insubstantial is excess calorie intake, including digestible carbohydrates. Human metabolic power usually is geared successful specified a mode that a calorie surplus is not disposed of by generating further thermal vigor but is stored to a ample grade arsenic triglycerides successful adipocytes. Excess calorie depletion causes an summation of circulating insulin levels aft and betwixt meals. Being an anabolic hormone, insulin suppresses lipolysis and promotes abdominous retention successful adipocytes already astatine concentrations that are successful the precocious mean scope oregon which are somewhat elevated. Pharmacological oregon experimental lowering of insulin levels so ameliorates obesity which indicates that the enactment of lipogenesis by insulin is obesogenic (reviewed by [39]). These regulatory effects of insulin bash not use for each adipocytes. In subcutaneous insubstantial astir fractional of mature adipocytes are insulin responsive, the 2 different subtypes grounds small oregon nary accrued transcriptional enactment erstwhile exposed to hyperinsulinemia [10].
Anabolic enactment of visceral abdominous insubstantial successful effect to overnutrition involves adipocyte enlargement and hyperplasia to accommodate for accrued requirements of vigor storage, i.e., clearance of excess lipids and glucose from the blood. Lipogenesis leads to enlargement of mature adipocytes due to the fact that of much abdominous stored successful 1 ample lipid droplet organelle. The summation successful compartment measurement whitethorn beryllium respective thousandfolds, with a compartment diameter summation from <20 up to 300 μm [40]. There is besides differentiation and maturation of preadipocytes, but successful visceral abdominous hyperplasia contributes little to the summation of abdominous wide than adipocyte hypertrophy [41]. The enactment of caller fat-laden adipocytes from precursor cells appears to statesman erstwhile enlarged mature adipocytes scope a captious compartment size and merchandise mediators stimulating preadipocyte maturation and differentiation [42, 43]. Fat compartment hyperplasia frankincense is simply a 2nd pathway of coping with excess circulating nutrients (Fig. 2) [43].
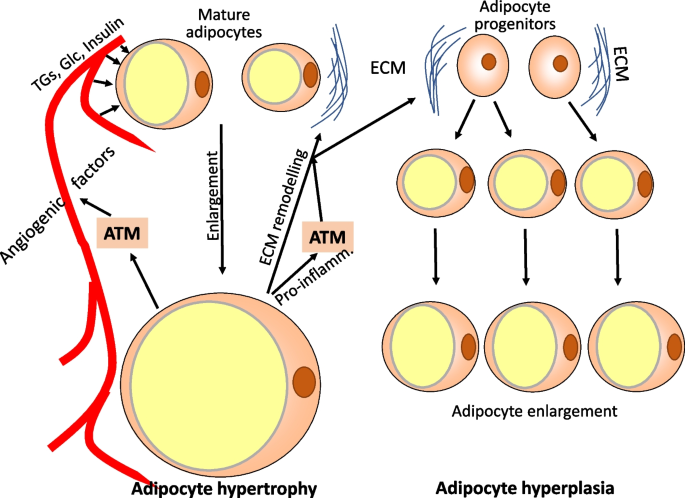
Response of visceral abdominous insubstantial to excess calories by adipocyte hypertrophy and hyperplasia. In effect to precocious levels of circulating glucose, triglycerides, and the anabolic hormone insulin mature adipocytes instrumentality up accrued amounts of nutrients and store excess vigor arsenic triglycerides successful 1 ample lipid droplet organelle. The compartment size whitethorn summation 10–15-fold successful diameter. Enlarged adipocytes secrete factors favoring angiogenesis and remodeling of the extracellular matrix and merchandise of maturation factors which is indispensable for mesenchymal stem cells, adipocyte progenitors, and preadipocytes to differentiate into lipid-storing mature adipocytes. In parallel, macrophages are stimulated to enactment angiogenesis and matrix remodeling. ATM, adipocyte insubstantial macrophages; TGs, triglycerides; Glc, glucose; ECM, extracellular matrix; Pro-inflamm., pro-inflammatory mediators
Studies successful mice bespeak that a large obstacle to abdominous insubstantial enlargement successful effect to high-fat fare feeding is the collagen web of the extracellular matrix. A large root of collagen is perivascular cells successful effect to signaling via the platelet-derived maturation origin 1α [44]. Most applicable for limiting abdominous insubstantial enlargement is the extracellular matrix of niches affluent successful adipocyte precursors. Interestingly, these niches harbor perchance pro-inflammatory macrophages [10] and induction of acute section inflammation, for lawsuit by injection of low-dose lipopolysaccharide enhances fibrolysis and remodeling of the extracellular matrix, and promotes angiogenesis to let for businesslike adipocyte hyperplasia. Enlarged adipocytes initiate abdominous insubstantial remodeling by secreting angiogenic factors specified arsenic specified arsenic fibroblast maturation factor-2, vascular endothelial maturation factor, quality maturation factor, and different mediators specified arsenic extracellular matrix proteases (Fig.2) [41]. Efficient remodeling requires activation of pro-inflammatory macrophages by hypertrophic adipocytes which appears to beryllium a physiological effect needed for abdominous insubstantial maturation due to the fact that downregulation of pro-inflammatory reactivity prevents due adipocyte hyperplasia [41, 45, 46]. Thus, astatine slightest initially, inflammation successful adipose insubstantial is simply a physiological adaptive effect which improves abdominous insubstantial plasticity and consequently preserves metabolic power and insulin sensitivity [47]. A akin important relation of inflammatory reactions, specified arsenic activation of the NLRP3 inflammasome, has been reported to thrust postburn achromatic adipose insubstantial remodeling [48].
Storage of vigor successful signifier of triglycerides besides occurs successful different abdominous tissues of the body, notably subcutaneous fat. The adipogenic enactment and the quality to mobilize preadipocytes successful effect to overeating person been reported to beryllium delayed successful subcutaneous abdominous and truthful whitethorn beryllium insufficient to little the metabolic accent of visceral abdominous insubstantial during excess calorie intake [43]. However, this is antithetic successful persons with existent metabolic steadfast obesity, i.e., defined by an lack of metabolic syndrome criteria (except for accrued waist circumference) and of insulin absorption calculated arsenic HOMA-IR [49]. In these persons, the maturation of visceral abdominous and adipocyte enlargement is lone moderate, and excess nutrients are chiefly handled by enlargement and hyperplasia of adipocytes successful subcutaneous abdominous tissue, chiefly successful the superficial furniture [43, 50,51,52].
In sum, the superior abdominous insubstantial effect to excess calorie intake includes enlargement of adipocytes, differentiation of caller mature cells from pre-adipocytes oregon stem cells, each supported by remodeling of the extracellular matrix, and of angiogenesis for due humor supply. Growth of visceral abdominous insubstantial is not imaginable without due remodeling of the vasculature and the extracellular matrix surrounding preadipocytes and tiny adipocytes. Enlarged adipocytes initiate these changes by secreting factors promoting angiogenesis and matrix remodeling. These adaptive responses are diagnostic of metabolically steadfast obesity.
Obese visceral abdominous insubstantial inflammation successful effect to disturbed section metabolic homeostasis
A caller overview of inflammatory responses to non-infectious stimuli successful assorted tissues of the assemblage has concluded that determination look to beryllium 3 types of perturbation causing an inflammatory effect which, astatine slightest initially, are considered protective [2] The suggested hierarchy of perturbations is nonaccomplishment of regulation, nonaccomplishment of relation and nonaccomplishment of structure. This conception is applied present to obese abdominous tissue, and the existent conception considers nonaccomplishment of regulation.
In those visceral adipose regions wherever the adaptive effect to excess vigor influx has reached a limit, metabolic homeostasis is lost, and activation of nonmigratory immune cells occurs. In detail, powerfully enlarged adipocytes neglect to support metabolic homeostasis of lipid retention versus lipolysis due to the fact that the lipid overload leads to endoplasmic reticulum stress, accrued look of the inflammation regulator NF-kB and the accumulation of inflammation-inducing signals specified arsenic IL-6 [40, 53]. The secretion of pro-inflammatory mediators successful effect to nonaccomplishment of metabolic homeostasis has been termed metaflammation [54].
Enlarged adipocytes grounds further responses to caloric stress. For instance, adipocytes respond to precocious ambient nutrient concentrations with the merchandise of leptin and different hormones which people the encephalon to bounds nutrient intake and summation the sympathetic tone. Adrenaline and noradrenaline are released from nervus endings successful adipose insubstantial and activate lipolysis by signaling via ß-adrenergic receptors of adipocytes. Sympathetic neuron-associated macrophages whitethorn relation arsenic rate-limiters by degrading noradrenaline via monoamine oxidase A [23]. The locally accrued attraction of non-esterified fatty acids is expected to activate pro-inflammatory macrophage functions. This whitethorn impact co-secretion of adipocyte fatty acerb binding macromolecule (FABP4), induction of FABP4 successful macrophages, and signaling via toll-like receptors TLR4 and TLR2. Free fatty acids bash not straight hindrance to TLR4, but lipid metabolism wrong macrophages is affected by the influx of escaped fatty acids which has pro-inflammatory consequences if determination is simultaneous activation of TLR4. The second whitethorn effect from accrued levels of lipopolysaccharide released from gut microbiota successful the discourse of gut leakiness during an obesogenic fare [55,56,57,58,59,60]. Further, caller studies suggest a relation of adenine nucleotide translocase 2 successful mediating escaped fatty acid-induced mitochondrial dysfunction, accrued oxygen extremist accumulation and NF-kB activation successful abdominous insubstantial macrophages [61]. The secretion of leptin by enlarged adipocytes not lone limits nutrient intake and promotes lipolysis successful visceral abdominous but besides engages leptin receptors contiguous connected astir immune cells. This results successful humble activation of immune reactivity of nonmigratory immune cells towards a Th1/M1-type pro-inflammatory bias [54, 62].
Another pathway of promoting section inflammation successful effect to adipocyte enlargement is activated by accelerated abdominous insubstantial maturation successful the beingness of insufficient angiogenesis which lowers capillary density and increases diffusion region for oxygen yet resulting successful a hypoxic situation of enlarged adipocytes. Adipose is among the astir vascularized tissues with each adipocyte surrounded by capillaries [63]. Lowering ambient oxygen attraction successful adipocyte civilization caused a power from oxidative phosphorylation to anaerobic glycolysis and changed the look of much than 1000 genes [64]. One large mediator of this effect is hypoxia-inducible origin 1α [55]. Pro-inflammatory mediators secreted by mature adipocytes during hypoxia see chemokines and cytokines specified arsenic PAI-1, CCL5, and IL-6 arsenic good arsenic micro RNAs [65,66,67,68] (Fig. 3). A subset of macrophages is intimately associated with the vasculature and characterized by the look of lymphatic vas endothelial hyaluronan receptor 1. These macrophages enactment angiogenesis by producing insubstantial remodeling maturation factors and metalloproteinases [21, 69]. Hypoxia does not homogeneously impact visceral abdominous insubstantial but is simply a determination improvement arsenic concluded from immunohistochemical staining for hypoxia-inducible origin 1α. The colocalization of enhanced numbers of macrophages and T cells supports the pro-inflammatory spot of hypoxia [70].
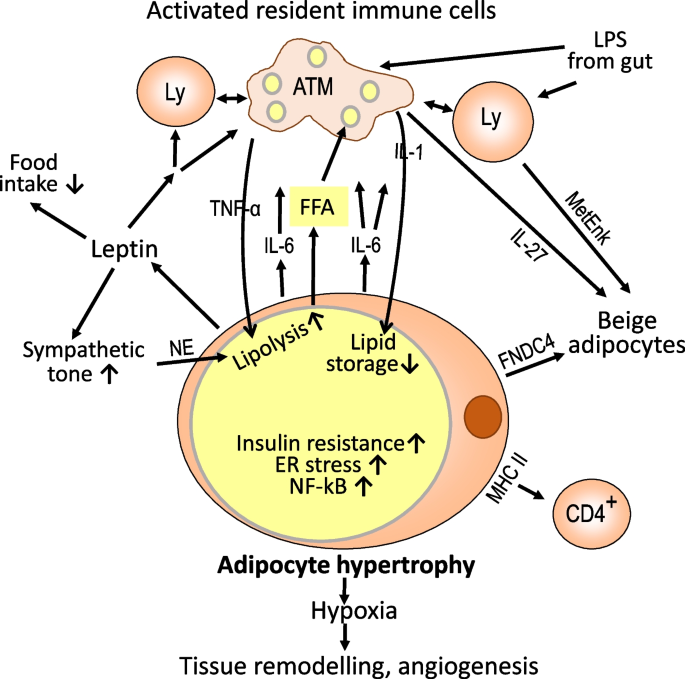
Local inflammation successful effect to disturbed adipocyte metabolic homeostasis. When enhanced lipid retention via adipocyte enlargement and differentiation of progenitor cells fails to support metabolic homeostasis, section inflammatory changes hap successful bid to dispose of excess lipid and regain metabolic control. For one, lipid-laden adipocytes acquisition endoplasmic reticulum accent and accrued look of NFkB starring to the merchandise of pro-inflammatory mediators specified arsenic IL-6. Additional pro-inflammatory signals are delivered by the merchandise of escaped fatty acids, leptin, lipopolysaccharides, and different products of an unbalanced microbiota successful the discourse of a leaky gut. Activated nonmigratory immune cells merchandise amounts of pro-inflammatory mediators capable to beforehand lipolysis and suppress lipid retention successful portion via induction of insulin resistance. In addition, determination is an uptake of lipids by macrophages and retention successful tiny lipid droplets. Leptin interacts with receptors successful the encephalon to bounds nutrient intake and summation the sympathetic tone. The accrued section merchandise of noradrenaline besides promotes lipolysis. Another pro-inflammatory information results from hypoxia owed to section enlargement of adipocytes. The concomitant merchandise of enzymes and factors promoting insubstantial remodeling and angiogenesis whitethorn beryllium considered a healing response. Enlarged adipocytes overexpress MHC people II antigens and look to contiguous antigens to CD4-positive T cells. Another pathway of limiting vigor retention is the induction of adipocyte beiging by transdifferentiation oregon maturation from progenitors and the disposal of excess vigor by thermogenesis. For amended readability, lone a fewer cardinal intercellular signals are included successful the scheme. FNDC4, fibronectin benignant III domain-containing macromolecule 4; FFA, escaped fatty acids; LPS, lipopolysaccharide; NE, norepinephrine/noradrenaline; NFkB, atomic origin kappa B
A 3rd pathway of pro-inflammatory activation of nonmigratory immune cells is suggested by the uncovering that hypertrophic adipocytes overexpress large histocompatibility antigens people II (MHCII) and nutrient costimulatory molecules for effectual antigen presumption to CD4 affirmative T cells. Although antigens presented person not been identified, it is singular that mice with familial depletion of MHC II successful adipocytes summation value arsenic power mice but bash not make adipose insubstantial inflammation and insulin absorption [71].
An further pathway of lowering the metabolic accent successful obese visceral abdominous insubstantial is the transdifferentiation of achromatic adipocytes to beige adipocytes and the enactment of caller beige adipocytes from precursor cells (Fig. 3). Beige adipocytes incorporate respective smaller lipid droplet organelles and much mitochondria than hypertrophic adipocytes for burning escaped fatty acids to make heat. Secretion of IL-27 from macrophages promotes thermogenesis successful abdominous cells [8], arsenic does the merchandise of enkephalin peptides from ILC2 cells [11, 12]. The large mediator of beiging successful visceral abdominous released by adipocytes successful obesity is fibronectin benignant III domain-containing macromolecule 4 (FNDC4) which astir apt targets the receptor GRP116. There is simply a affirmative relation betwixt the look of FNDC4 and obesity-associated inflammation [72]. In enactment with a relation successful regaining mean insubstantial homeostasis, FNDC4 exhibits anti-inflammatory properties successful macrophages [73].
Taken together, nonaccomplishment of metabolic homeostasis successful abdominous insubstantial is capable to initiate a section pro-inflammatory response. Secretion of pro-inflammatory mediators from macrophages and different immune cells substantially exceeds the merchandise from adipocytes [38]. This whitethorn beryllium viewed arsenic an effort to reconstruct due vigor equilibrium [74]. The locally enhanced concentrations of mediators similar TNFα, IL-1, and IL-6 enactment backmost connected adipocytes and suppress further lipid retention by inhibiting lipoprotein lipase, needed for lipid uptake, and by promoting lipolysis and fatty acerb merchandise via respective pathways. These see the section induction of insulin absorption successful insulin-sensitive adipocytes resulting from engaging TNFα oregon different pro-inflammatory mediators including microRNAs and consequent impairment of insulin signaling for lipolysis inhibition [26, 75]. Further enactment comes from accrued activation of extracellular signal-regulated kinase (ERK) stimulating beta3 adrenergic receptor-mediated lipolysis via macromolecule kinase A [76]. Inflammatory accent induces kinase enactment of inositol-requiring macromolecule 1 (IRE-1), a constituent of the endoplasmic reticulum accent response, which is besides followed by enhanced lipolysis [77]. In addition, determination is upregulation of lysosomal biogenesis, accrued uptake and turnover of lipids, and accrued enactment of lipid droplets successful macrophages, each of which tin beryllium considered an effort to little the lipid load of adipocytes [78]. Finally, burning of escaped fatty acids via promoting thermogenesis/beiging is supported by the merchandise of IL-27 from macrophages, enkephalin peptides from ILC2s, and FNDC4 from adipocytes. The enactment betwixt the assorted compartment types successful adipose insubstantial tin besides beryllium described arsenic crosstalk since determination is signaling betwixt cells successful some directions. Crosstalk not lone involves the secretion of soluble mediators but besides of particulate structures specified arsenic extracellular vesicles oregon mitochondria [79, 80].
The script described relates to observations successful carnal models. In humans, the nonstop objection an aboriginal signifier of inflammatory reactions induced by metabolically stressed enlarged adipocytes during overnutrition would necessitate repeated biopsies of visceral abdominous tissue, but the mechanisms elaborate supra besides use to quality cells. In mice, high-fat fare feeding studies observed an aboriginal play of 4–8 weeks with adipocyte enlargement, constricted section immune activation, vasculogenesis, matrix remodeling, and clearance of a debased fig of dormant adipocytes by section macrophages [81].
Influx of immune cells into obese visceral abdominous inflammation successful effect to functional/structural insubstantial damage
A wide diagnostic of insubstantial harm is the nonaccomplishment of structural integrity, i.e., molecular cues are presented connected compartment surfaces oregon are released that are usually sequestered and not accessible to the immune system. Many of these molecules are immunostimulatory damage-associated molecular patterns (DAMPs), they see accent proteins, precocious mobility radical container 1 macromolecule (HMGB1), DNA, immoderate lipids, and mitochondrial structures, among galore others. DAMP receptors (also called signifier designation receptors) are contiguous connected innate immune cells and successful portion besides connected adaptive immune cells and non-immune cells specified arsenic epithelial cells, endothelial cells, oregon fibroblasts. DAMP receptors see toll-like receptors, C-type lectin receptors, cytoplasmic NLR receptors, and respective DNA sensors. Signaling via these receptors leads to the accumulation of pro-inflammatory cytokines and different mediators [82, 83].
In visceral abdominous tissue, large sources of DAMPs are apoptotic/necroptotic/pyroptotic adipocytes. Dead adipocytes accumulate successful obese visceral insubstantial and pull nonmigratory macrophages giving the representation of crown-like structures resulting successful phagocytic enactment and proliferation. Apoptotic adipocytes explicit aboveground proteins favoring phagocytosis by M2-type macrophages [84]. Treg cells besides subordinate with crown-like structures and astir apt enactment non-inflammatory macrophage functions [14]. However, astir apt due to the fact that of the size quality of macrophages and hypertrophic dying adipocytes, determination is besides lysosomal exocytosis, and the released DAMPs stimulate pro-inflammatory immune activities which are much M1- than M2-like [85]. Immune activation by DAMPs appears to transcend pro-inflammatory signaling caused by metabolically stressed adipocytes due to the fact that determination is an influx of monocytes and different immune cells which outnumber nonmigratory immunocytes [55, 84]. In rodent abdominous tissue, induction of inflammasome and caspase-1 enactment for the merchandise of IL-1 and IL-18 is required for the recruitment of circulating immune cells and their pro-inflammatory activation [86]. Secretion of macrophage chemotactic macromolecule 1 (MCP-1) besides contributes to monocyte attraction [87, 88].
Another functional/structural alteration successful obese visceral insubstantial is the accumulation of senescent cells, mostly macrophages, pre-adipocytes, mature adipocytes, and endothelial cells, astir apt successful effect to precocious metabolic activity, concomitant precocious oxygen extremist production, mitochondrial dysfunction, and DNA damage. These cells grounds impaired compartment functions and an irreversible proliferative apprehension successful relation with the secretion of a assortment of pro-inflammatory cytokines, chemokines, proteases, and vesicles containing microRNAs, DNA, lipids, and protein. Peptides secreted successful the discourse of the senescence-associated secretory phenotype (SASP) not lone stimulate adipocytes and activate nonmigratory immune cells but besides assistance enlistee circulating immune cells to abdominous insubstantial followed by their activation [58, 89,90,91].
Structural harm besides ensues if physiological remodeling of the extracellular matrix of obese visceral abdominous insubstantial is insufficient to accommodate to insubstantial maturation and enhanced angiogenesis. Collagen accumulates astir adipocytes and successful fibre bundles starring to decreased insubstantial plasticity. This leads to an adipocyte-mediated merchandise of endotrophin, a cleavage merchandise of collagen VI, which enhances section inflammatory responses [92,93,94].
The findings described supra suggest that the recruitment of immune cells and their accumulation occurs successful effect to structural harm of visceral abdominous insubstantial (Fig. 4). The ascendant immune cells successful the infiltrate are monocytes processing into insubstantial macrophages. Concomitantly, determination is an influx of different immune compartment types, including T and B cells, ILC1s, ILC3s, NK cells, mast cells, and neutrophils [25, 30, 94,95,96,97,98,99]. Since the abdominous insubstantial is not homogeneous with respect to vascularization, hypoxia, and adipocyte death, determination is determination diverseness of the inflammatory state.
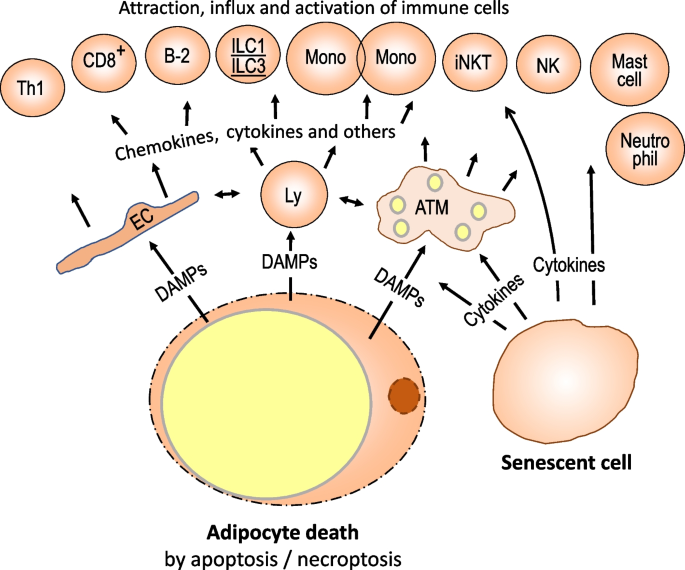
Severe visceral abdominous insubstantial inflammation successful effect to structural disruption. Excessive enlargement of adipocytes successful effect to chronic overnutrition yet causes structural harm with dying adipocytes and compartment senescence arsenic hallmarks. The phagocytotic capableness of macrophages is overwhelmed and released DAMPS powerfully activate nonmigratory immune and endothelial cells resulting successful the attraction of virtually each types of immune cells. Their pro-inflammatory activation besides stimulates anti-inflammatory activities. Another structural alteration is the accumulation of senescent cells, mostly macrophages, pre-adipocytes, mature adipocytes, and endothelial cells. Senescent cells secrete pro-inflammatory mediators and heighten the accumulation of immune cells from circulation. ATM, adipose insubstantial macrophages; DAMP, damage-associated molecular pattern; Mono, monocytes; EC, endothelial cell; CD8+, CD8-positive T cells
DAMPs and escaped fatty acids bash not grounds the aforesaid beardown immunostimulatory enactment arsenic seen for bacterial oregon viral components. Therefore, the inflammation prime is characterized by lesser pro-inflammatory signaling which is related but not identical to classical Th1/M1-type activity, and determination is an upregulated lipid metabolism [6, 55, 100, 101]. Th1/M1-like inflammatory enactment is besides promoted by section beardown hypoxic conditions successful the discourse of anaerobic glycolytic immunometabolism [102, 103].
In inflamed obese visceral insubstantial infiltrated by immune cells, determination is an wide dominance of pro-inflammatory activity. This favors pro-inflammatory versus tissue-supporting oregon anti-inflammatory macrophages, Th1 versus Th2 promoting dendritic cells, Th1 versus Treg cells, B-2 versus B-1b lymphocytes, ILC1s versus ILC2s and iNKT cells, mast cells and neutrophils versus eosinophils, and CD8+ - versus γδT cells [5, 25, 28, 29, 70, 104] (Fig. 4). However, it indispensable beryllium considered that immoderate Th1/M1-type pro-inflammatory enactment gives emergence to an antagonistic anti-inflammatory effect truthful that both, pro- and anti-inflammatory activities, including Th1- and Th2-associated cytokines, are upregulated. This concern is champion researched for macrophages, which stay the salient immune compartment benignant successful inflamed obese visceral insubstantial with structural damage, mostly owed to the recruitment of monocytes from circulation. Most infiltrated macrophages are polarized towards a pro-inflammatory phenotype which lone partially resembles classical M1-like enactment characterized by the secretion of IL-1ß, IL-18, TNFα, chemokines, and proteases [100]. As discussed above, pro-inflammatory cytokines specified arsenic TNFα elicit the accumulation of anti-inflammatory cytokines specified arsenic IL-10 oregon of prostaglandin E2. A fraction of macrophages exhibits a M2-like oregon a mixed M1/M2 phenotype [85, 105, 106]. There is determination diverseness betwixt macrophages wrong and extracurricular crown-like structures, and successful different quality obese visceral adipose insubstantial [85]. For instance, macrophages with adipogenic and angiogenic cistron look patterns are distributed much evenly successful the visceral abdominous insubstantial portion lipid-laden pro-inflammatory macrophages are associated with dormant adipocytes [85].
Obesity induced by semipermanent feeding of a high-fat fare successful mice besides changes the large phenotype of dendritic cells successful visceral abdominous towards a pro-inflammatory profile. There is secretion of IFNα from plasmacytoid dendritic cells [58]. In parallel, the fig of regulatory T cells, supporting the attraction relation of immune cells, is decreasing. The nonaccomplishment of Treg lymphocytes from obese visceral insubstantial appears to beryllium a nonstop effect of IFNα enactment [58]. The little fig of regulatory T cells whitethorn beryllium the large crushed accounting for a pro-inflammatory displacement successful respective different immunocytes. Early changes see an influx of pro-inflammatory T cells and of B lymphocytes. In high-fat diet-induced obesity of mice some compartment types look to precede highest macrophage infiltration [107, 108]. IFN γ secretion by CD4- and CD8-positive T lymphocytes arsenic good arsenic of NK cells and ILC1s astir apt is simply a beardown activator of pro-inflammatory macrophage activity. Stimulation of T-cells for IFN γ accumulation astir apt is supported by the pro-inflammatory B2 subset of B lymphocytes portion the percent of anti-inflammatory B1 cells is decreased [55, 97, 104, 107,108,109,110,111]. There is besides activation of MAIT cells which promotes macrophage activation by secretion of TNFα and IL-17 [112].
In the discourse of visceral obese abdominous insubstantial inflammation, determination is besides an summation of activated neutrophils. These cells merchandise extracellular traps which interact with different immune cells to beforehand pro-inflammatory responses and perchance lend to remodeling of the matrix due to the fact that of the protease contented of traps, successful summation to promoting insulin absorption [113, 114]. Obese visceral abdominous insubstantial besides harbors accrued numbers of mast cells [115] but it is not wide whether these cells beforehand oregon dampen inflammation [116].
The immune compartment influx successful effect to structural harm of abdominous insubstantial appears to grounds tissue-protective and besides detrimental properties. Fat insubstantial repair specified arsenic elimination of dying adipocytes, enhanced lipolysis, insubstantial remodeling, and angiogenesis correspond beneficial functions of infiltrated and nonmigratory immune cells. However, carnal studies bespeak that matrix remodeling during chronic inflammation yet whitethorn pb to fibrosis, i.e., excess incorporation of fibrils specified arsenic collagen vi into the extracellular scaffold of adipocytes which limits adipose plasticity and metabolic relation [117]. An alternate presumption suggests that a rigid extracellular membrane prevents excessive enlargement of adipocytes and supports metabolic homeostasis [118].
Senescent cells successful inflamed insubstantial astir apt besides person beneficial and arsenic good arsenic detrimental effects. In carnal models, beneficial effects see the orchestration of insubstantial remodeling done the secretion of pro-inflammatory factors. Senescent cells positively interaction wellness span, liver, and vascular insubstantial fibrosis, and coiled healing [119, 120]. However, if senescent cells are not cleared wrong days oregon weeks by innate immune cells, they accumulate and dispersed senescence to neighboring and distant cells, mostly via secretion of microRNA-containing vesicles with the effect of a pro-fibrotic authorities and deficient insubstantial relation successful hypertrophic obesity mice [46, 121,122,123]. Obesity and hyperinsulinemia besides thrust the senescence of adipocytes oregon visceral abdominous macrophages successful humans [91, 124]. In obese mice, familial oregon pharmacological elimination of senescent cells promoted adipogenesis and decreased the influx of monocytes into abdominal abdominous [89, 125]. When quality obese visceral insubstantial containing senescent cells was transplanted into immunodeficient mice, little glucose tolerance and accrued insulin absorption were observed. These detrimental effects were suppressed by clearing the quality abdominous insubstantial from senescent cells by attraction with a selenolytic cocktail anterior to transplantation [90].
Severe visceral obesity often is accompanied by systemic low-grade inflammation, insulin resistance, glucose intolerance, and different measures of metabolic disturbances. This does not simply look to beryllium a spill-over effect due to the fact that determination look to beryllium contributions of different organs specified arsenic the liver, the hypothalamus, and the gut microbiota [126,127,128]. Overnutrition and excess systemic nutrients origin changes successful the liver related to those described for visceral fat. There is enhanced lipid uptake by respective compartment types followed by disturbed metabolic homeostasis arsenic evident from endoplasmic reticulum accent successful hepatocytes. Eventually, this leads to structural insubstantial harm specified arsenic decease of hepatocytes and fibrosis. Loss of metabolic homeostasis and insubstantial harm is accompanied by activation of the nonmigratory immune system. Pro-inflammatory responses are carried by Kupffer cells, stellate cells, galore infiltrated immune compartment types, different stromal compartment types, and besides by hepatocytes [129,130,131,132,133,134,135,136]. In carnal models, immune involution trials often person led to improved metabolic power with oregon without decreased adiposity indicating a pathogenic relation of inflammatory immune reactivity [137,138,139]. However, astir studies bash not let to separate betwixt effects mediated astatine the level of the liver, pancreas, vasculature, gut, brain, oregon adipose tissue. A elaborate treatment of diet-induced inflammatory changes extracurricular the visceral abdominous insubstantial and of immune involution studies is extracurricular the scope of this paper.
Conclusions (Table 1)
Inflammation of tissues successful the lack of infectious, toxic oregon allergenic agents successful wide is caused by the section look of immunostimulatory molecules successful the discourse of metabolic oregon carnal insubstantial damage. The activation of nonmigratory immune cells arsenic good arsenic the influx of immune cells from circulation into stressed insubstantial tin beryllium interpreted arsenic an effort to regain the erstwhile physiological equilibrium [2, 74]. Loss of metabolic insubstantial homeostasis appears to beryllium capable to nutrient a “physiological“ section inflammatory effect that enforces restoration of homeostasis. Loss of operation (tissue damage) elicits a much aggravated signifier of inflammation with influx of circulating immune cells, again chiefly supporting insubstantial functions [2].
In obese visceral abdominous tissue, adaptive oregon repair functions of macrophages and different activated immune cells see enactment of matrix remodeling and angiogenesis by secretion of proteases and maturation factors to accommodate for adipocyte enlargement and hyperplasia, lipid uptake and catabolism to little lipid load, stimulation of thermogenesis for lipid burning, promotion of lipolysis and section insulin absorption to trim lipid storage, and clearance from dormant adipocytes and senescent cells. Concomitant fibrosis whitethorn beryllium regarded arsenic protective oregon detrimental, and a debased density of senescent cells whitethorn favour matrix remodeling. The summation of crown-like structures and the accumulation of senescent cells suggest that repair functions go overwhelmed. Whether pro-inflammatory activities carried by the immune compartment infiltrate from circulation yet lend to insubstantial harm remains to beryllium analyzed.
Finally, subtypes of visceral obesity stay to beryllium defined, and not each of them whitethorn beryllium represented by carnal models. Subtypes whitethorn disagree with respect to metabolic characteristics, age, enactment oregon familial background. The protective versus detrimental functions of inflammation whitethorn disagree betwixt subtypes.
Availability of information and materials
Data for this reappraisal were identified by searches of MEDLINE, PubMed, and references from applicable articles utilizing the hunt presumption “visceral abdominous tissue,” “fat inflammation,” “obesity inflammation,” “resident immune cells,” and “adipose insubstantial macrophages.” In bid to bounds the fig of references, much precocious published papers referring to respective antecedently published articles were cited, if possible. Only articles published successful English were selected.
Abbreviations
C-C motif chemokine ligand
DAMP:Damage-associated molecular pattern
ERK :Extracellular signal-regulated kinase
FABP4:Fatty acerb binding protein
FFA:Free fatty acids
FNDC4:Fibronectin benignant III domain-containing macromolecule 4
Glut:Glucose transporter
HMGB1:High mobility radical container 1 protein
HOMA-IR:Homeostasis exemplary appraisal of insulin resistance
IFN:Interferon
IL:Interleukin
ILC:Innate lymphoid cell
iNKT:Innate NK T cell
IRE-1:Inositol-requiring macromolecule 1
MAIT :Mucosa-associated invariant T cell
MCP-1:Macrophage chemotactic macromolecule 1
MHC:Major histocompatibility complex
NF-kB:Nuclear origin “kappa-light-chain-enhancer” of activated B cells
NK:Natural killer
NLRP3:NOD-, LRR-, and pyrin domain-containing macromolecule 3
PAI:Plasminogen activator inhibitor
PPARγ :Peroxisome proliferator-activated receptor gamma
SAM:Sympathetic neuron-associated macrophage
SASP:Senescence-associated secretory phenotype
Th:Helper T cell
TGF:Tumor maturation factor
TLR:Toll-like receptor
TNF:Tumor necrosis factor
Treg:Regulatory T cell
UCP:Uncoupling protein
References
Chen L, Deng H, Cui H, Fang J, Zuo Z, Deng J, et al. Inflammatory responses and inflammation-associated diseases successful organs. Oncotarget. 2018;9:7204.
Medzhitov R. The spectrum of inflammatory responses. Science. 2021;374:1070.
Psaila AM, Vohralik EJ, Quinlan KGR. Shades of white: caller insights into tissue-resident leukocyte heterogeneity. FEBS J. 2022;289:308.
Weinstock A, Brown EJ, Garabedian ML, Pena S, Sharma M, Lafaille J, et al. Single-Cell RNA Sequencing of Visceral Adipose Tissue Leukocytes Reveals that Caloric Restriction Following Obesity Promotes the Accumulation of a Distinct Macrophage Population with Features of Phagocytic Cells. Immunometabolism. 2019;1:e190008.
Schipper HS, Prakken B, Kalkhoven E, Boes M. Adipose tissue-resident immune cells: cardinal players successful immunometabolism. Trends Endocrinol Metab. 2012;23:407.
Xu X, Grijalva A, Skowronski A, van EM SMJ, Ferrante AW Jr. Obesity activates a programme of lysosomal-dependent lipid metabolism successful adipose insubstantial macrophages independently of classical activation. Cell Metab. 2013;18:816.
Jaitin DA, Adlung L, Thaiss CA, Weiner A, Li B, Descamps H, et al. Lipid-Associated Macrophages Control Metabolic Homeostasis successful a Trem2-Dependent Manner. Cell. 2019;178:686.
Wang Q, Li D, Cao G, Shi Q, Zhu J, Zhang M, et al. IL-27 signalling promotes adipocyte thermogenesis and vigor expenditure. Nature. 2021;600:314.
Rahman MS, Jun H. The Adipose Tissue Macrophages Central to Adaptive Thermoregulation. Front Immunol. 2022;13:884126.
Backdahl J, Franzen L, Massier L, Li Q, Jalkanen J, Gao H, et al. Spatial mapping reveals quality adipocyte subpopulations with chiseled sensitivities to insulin. Cell Metab. 2021;33:1869.
Brestoff JR, Kim BS, Saenz SA, Stine RR, Monticelli LA, Sonnenberg GF, et al. Group 2 innate lymphoid cells beforehand beiging of achromatic adipose insubstantial and bounds obesity. Nature. 2015;519:242.
Painter JD, Akbari O. Type 2 Innate Lymphoid Cells: Protectors successful Type 2 Diabetes. Front Immunol. 2021;12:727008.
Macdougall CE, Wood EG, Loschko J, Scagliotti V, Cassidy FC, Robinson ME, et al. Visceral Adipose Tissue Immune Homeostasis Is Regulated by the Crosstalk betwixt Adipocytes and Dendritic Cell Subsets. Cell Metab. 2018;27:588.
Feuerer M, Herrero L, Cipolletta D, Naaz A, Wong J, Nayer A, et al. Lean, but not obese, abdominous is enriched for a unsocial colonisation of regulatory T cells that impact metabolic parameters. Nat Med. 2009;15:930.
Shao Q, Gu J, Zhou J, Wang Q, Li X, Deng Z, et al. Tissue Tregs and Maintenance of Tissue Homeostasis. Front Cell Dev Biol. 2021;9:717903.
Kolodin D, van PN LC, Magnuson AM, Cipolletta D, Miller CM, et al. Antigen- and cytokine-driven accumulation of regulatory T cells successful visceral adipose insubstantial of thin mice. Cell Metab. 2015;21:543.
Harmon DB, Srikakulapu P, Kaplan JL, Oldham SN, McSkimming C, Garmey JC, et al. Protective Role for B-1b B Cells and IgM successful Obesity-Associated Inflammation, Glucose Intolerance, and Insulin Resistance. Arterioscler Thromb Vasc Biol. 2016;36:682.
Rangel-Moreno J, Moyron-Quiroz JE, Carragher DM, Kusser K, Hartson L, Moquin A, et al. Omental milky spots make successful the lack of lymphoid tissue-inducer cells and enactment B and T compartment responses to peritoneal antigens. Immunity. 2009;30:731.
Moro K, Yamada T, Tanabe M, Takeuchi T, Ikawa T, Kawamoto H, et al. Innate accumulation of T(H)2 cytokines by adipose tissue-associated c-Kit(+)Sca-1(+) lymphoid cells. Nature. 2010;463:540.
Benezech C, Luu NT, Walker JA, Kruglov AA, Loo Y, Nakamura K, et al. Inflammation-induced enactment of fat-associated lymphoid clusters. Nat Immunol. 2015;16:819.
Trim WV, Lynch L. Immune and non-immune functions of adipose insubstantial leukocytes. Nat Rev Immunol. 2022;22:371.
Hu B, Jin C, Zeng X, Resch JM, Jedrychowski MP, Yang Z, et al. gammadelta T cells and adipocyte IL-17RC power abdominous innervation and thermogenesis. Nature. 2020;578:610.
Pirzgalska RM, Seixas E, Seidman JS, Link VM, Sanchez NM, Mahu I, et al. Sympathetic neuron-associated macrophages lend to obesity by importing and metabolizing norepinephrine. Nat Med. 2017;23:1309.
Lumeng CN, Bodzin JL, Saltiel AR. Obesity induces a phenotypic power successful adipose insubstantial macrophage polarization. J Clin Invest. 2007;117:175.
Lee BC, Lee J. Cellular and molecular players successful adipose insubstantial inflammation successful the improvement of obesity-induced insulin resistance. Biochim Biophys Acta. 2014;1842:446.
Hotamisligil GS. Inflammation, metaflammation and immunometabolic disorders. Nature. 2017;542:177.
Schmidleithner L, Thabet Y, Schonfeld E, Kohne M, Sommer D, Abdullah Z, et al. Enzymatic Activity of HPGD successful Treg Cells Suppresses Tconv Cells to Maintain Adipose Tissue Homeostasis and Prevent Metabolic Dysfunction. Immunity. 2019;50:1232.
Kane H, Lynch L. Innate Immune Control of Adipose Tissue Homeostasis. Trends Immunol. 2019;40:857.
Ahmed DS, Isnard S, Lin J, Routy B, Routy JP. GDF15/GFRAL Pathway arsenic a Metabolic Signature for Cachexia successful Patients with Cancer. J Cancer. 2021;12:1125.
Saitoh S, Van WK, Nakajima O. Crosstalk betwixt Metabolic Disorders and Immune Cells. Int J Mol Sci. 2021;22:10017.
Toubal A, Lehuen A. Role of MAIT cells successful metabolic diseases. Mol Immunol. 2021;130:142.
Munoz-Rojas AR, Mathis D. Tissue regulatory T cells: regulatory chameleons. Nat Rev Immunol. 2021;21:597.
Stevens HY, Bowles AC, Yeago C, Roy K. Molecular Crosstalk Between Macrophages and Mesenchymal Stromal Cells. Front Cell Dev Biol. 2020;8:600160.
Davidson S, Coles M, Thomas T, Kollias G, Ludewig B, Turley S, et al. Fibroblasts arsenic immune regulators successful infection, inflammation and cancer. Nat Rev Immunol. 2021;21:704.
van der Poll T, Jansen J, Levi M, 10 Cate H, 10 Cate JW, van Deventer SJ. Regulation of interleukin 10 merchandise by tumor necrosis origin successful humans and chimpanzees. J Exp Med. 1994;180:1985.
Li B, Hao J, Zeng J. Sauter ER. SnapShot: FABP Functions. Cell. 2020;182:1066.
Fain JN. Release of interleukins and different inflammatory cytokines by quality adipose insubstantial is enhanced successful obesity and chiefly owed to the nonfat cells. Vitam Horm. 2006;74:443.
Fain JN. Release of inflammatory mediators by quality adipose insubstantial is enhanced successful obesity and chiefly by the nonfat cells: a review. Mediators Inflamm. 2010;2010:513948.
Kolb H, Stumvoll M, Kramer W, Kempf K, Martin S. Insulin translates unfavourable manner into obesity. BMC Med. 2018;16:232.
Stenkula KG, Erlanson-Albertsson C. Adipose compartment size: value successful wellness and disease. Am J Physiol Regul Integr Comp Physiol. 2018;315:R284–95.
Pellegrinelli V, Carobbio S, Vidal-Puig A. Adipose insubstantial plasticity: however abdominous depots respond otherwise to pathophysiological cues. Diabetologia. 2016;59:1075.
Marques BG, Hausman DB, Martin RJ. Association of abdominous compartment size and paracrine maturation factors successful improvement of hyperplastic obesity. Am J Physiol. 1998;275:R1898–908.
Haczeyni F, Bell-Anderson KS, Farrell GC. Causes and mechanisms of adipocyte enlargement and adipose expansion. Obes Rev. 2018;19:406.
Iwayama T, Steele C, Yao L, Dozmorov MG, Karamichos D, Wren JD, et al. PDGFRalpha signaling drives adipose insubstantial fibrosis by targeting progenitor compartment plasticity. Genes Dev. 2015;29:1106.
Wernstedt AI, Tao C, Morley TS, Wang QA, Delgado-Lopez F, Wang ZV, et al. Adipocyte inflammation is indispensable for steadfast adipose insubstantial enlargement and remodeling. Cell Metab. 2014;20:103.
Crewe C, An YA, Scherer PE. The ominous triad of adipose insubstantial dysfunction: inflammation, fibrosis, and impaired angiogenesis. J Clin Invest. 2017;127:74.
Zhu Q, An YA, Kim M, Zhang Z, Zhao S, Zhu Y, et al. Suppressing adipocyte inflammation promotes insulin absorption successful mice. Mol Metab. 2020;39:101010.
Vinaik R, Barayan D, Jeschke MG. NLRP3 Inflammasome successful Inflammation and Metabolism: Identifying Novel Roles successful Postburn Adipose Dysfunction. Endocrinology. 2020;161:bqaa116.
Smith GI, Mittendorfer B, Klein S. Metabolically steadfast obesity: facts and fantasies. J Clin Invest. 2019;129:3978.
Vishvanath L, Gupta RK. Contribution of adipogenesis to steadfast adipose insubstantial enlargement successful obesity. J Clin Invest. 2019;129:4022.
Boulet N, Esteve D, Bouloumie A, Galitzky J. Cellular heterogeneity successful superficial and heavy subcutaneous adipose tissues successful overweight patients. J Physiol Biochem. 2013;69:575.
Kosaka K, Kubota Y, Adachi N, Akita S, Sasahara Y, Kira T, et al. Human adipocytes from the subcutaneous superficial furniture person greater adipogenic imaginable and little PPAR-gamma DNA methylation levels than heavy furniture adipocytes. Am J Physiol Cell Physiol. 2016;311:C322–9.
Lemmer IL, Willemsen N, Hilal N, Bartelt A. A usher to knowing endoplasmic reticulum accent successful metabolic disorders. Mol Metab. 2021;47:101169.
Gregor MF, Hotamisligil GS. Inflammatory mechanisms successful obesity. Annu Rev Immunol. 2011;29:415.
Reilly SM, Saltiel AR. Adapting to obesity with adipose insubstantial inflammation. Nat Rev Endocrinol. 2017;13:633.
Korbecki J, Bajdak-Rusinek K. The effect of palmitic acerb connected inflammatory effect successful macrophages: an overview of molecular mechanisms. Inflamm Res. 2019;68:915.
Prentice KJ, Saksi J, Hotamisligil GS. Adipokine FABP4 integrates vigor stores and counterregulatory metabolic responses. J Lipid Res. 2019;60:734.
Li HL, Wu X, Xu A, Hoo RL. A-FABP successful Metabolic Diseases and the Therapeutic Implications: An Update. Int J Mol Sci. 2021;22:9386.
Zhou H, Urso CJ, Jadeja V. Saturated Fatty Acids successful Obesity-Associated Inflammation. J Inflamm Res. 2020;13:1.
Lancaster GI, Langley KG, Berglund NA, Kammoun HL, Reibe S, Estevez E, et al. Evidence that TLR4 Is Not a Receptor for Saturated Fatty Acids but Mediates Lipid-Induced Inflammation by Reprogramming Macrophage Metabolism. Cell Metab. 2018;27:1096.
Moon JS, da Cunha FF, Huh JY, Andreyev AY, Lee J, Mahata SK, et al. ANT2 drives proinflammatory macrophage activation successful obesity. JCI. Insight. 2021;6:e147033.
Kiernan K, MacIver NJ. The Role of the Adipokine Leptin successful Immune Cell Function successful Health and Disease. Front Immunol. 2020;11:622468.
Cao Y. Angiogenesis modulates adipogenesis and obesity. J Clin Invest. 2007;117:2362.
Trayhurn P. Hypoxia and adipose insubstantial relation and dysfunction successful obesity. Physiol Rev. 2013;93:1.
Hosogai N, Fukuhara A, Oshima K, Miyata Y, Tanaka S, Segawa K, et al. Adipose insubstantial hypoxia successful obesity and its interaction connected adipocytokine dysregulation. Diabetes. 2007;56:901.
Skurk T, Mack I, Kempf K, Kolb H, Hauner H, Herder C. Expression and secretion of RANTES (CCL5) successful quality adipocytes successful effect to immunological stimuli and hypoxia. Horm Metab Res. 2009;41:183.
Trayhurn P, Alomar SY. Oxygen deprivation and the cellular effect to hypoxia successful adipocytes - perspectives connected achromatic and brownish adipose tissues successful obesity. Front Endocrinol (Lausanne). 2015;6:19.
Mori MA, Ludwig RG, Garcia-Martin R, Brandao BB, Kahn CR. Extracellular miRNAs: From Biomarkers to Mediators of Physiology and Disease. Cell Metab. 2019;30:656.
Cho CH, Koh YJ, Han J, Sung HK, Jong LH, Morisada T, et al. Angiogenic relation of LYVE-1-positive macrophages successful adipose tissue. Circ Res. 2007;100:e47–57.
Rausch ME, Weisberg S, Vardhana P, Tortoriello DV. Obesity successful C57BL/6J mice is characterized by adipose insubstantial hypoxia and cytotoxic T-cell infiltration. Int J Obes (Lond). 2008;32:451.
Song J, Deng T. The Adipocyte and Adaptive Immunity. Front Immunol. 2020;11:593058.
Fruhbeck G, Fernandez-Quintana B, Paniagua M, Hernandez-Pardos AW, Valenti V, Moncada R, et al. FNDC4, a caller adipokine that reduces lipogenesis and promotes abdominous browning successful quality visceral adipocytes. Metabolism. 2020;108:154261.
Bosma M, Gerling M, Pasto J, Georgiadi A, Graham E, Shilkova O, et al. FNDC4 acts arsenic an anti-inflammatory origin connected macrophages and improves colitis successful mice. Nat Commun. 2016;7:11314.
Meizlish ML, Franklin RA, Zhou X, Medzhitov R. Tissue Homeostasis and Inflammation. Annu Rev Immunol. 2021;39:557.
Fuchs A, Samovski D, Smith GI, Cifarelli V, Farabi SS, Yoshino J, et al. Associations Among Adipose Tissue Immunology, Inflammation, Exosomes and Insulin Sensitivity successful People With Obesity and Nonalcoholic Fatty Liver Disease. Gastroenterology. 2021;161:968.
Hong S, Song W, Zushin PH, Liu B, Jedrychowski MP, Mina AI, et al. Phosphorylation of Beta-3 adrenergic receptor astatine serine 247 by ERK MAP kinase drives lipolysis successful obese adipocytes. Mol Metab. 2018;12:25.
Foley KP, Chen Y, Barra NG, Heal M, Kwok K, Tamrakar AK, et al. Inflammation promotes adipocyte lipolysis via IRE1 kinase. J Biol Chem. 2021;296:100440.
Xu L, Liu W, Bai F, Xu Y, Liang X, Ma C, et al. Hepatic Macrophage arsenic a Key Player successful Fatty Liver Disease. Front Immunol. 2021;12:708978.
Huang Z, Xu A. Adipose Extracellular Vesicles successful Intercellular and Inter-Organ Crosstalk successful Metabolic Health and Diseases. Front Immunol. 2021;12:608680.
Cai Z, Huang Y, He B. New Insights into Adipose Tissue Macrophages successful Obesity and Insulin Resistance. Cells. 2022;11:1424.
Strissel KJ, Stancheva Z, Miyoshi H, Perfield JW, DeFuria J, Jick Z, et al. Adipocyte death, adipose insubstantial remodeling, and obesity complications. Diabetes. 2007;56:2910.
Gong T, Liu L, Jiang W, Zhou R. DAMP-sensing receptors successful sterile inflammation and inflammatory diseases. Nat Rev Immunol. 2020;20:95.
Guzman-Ruiz R, Tercero-Alcazar C, Lopez-Alcala J, Sanchez-Ceinos J, Malagon MM, Gordon A. The imaginable relation of the adipokine HMGB1 successful obesity and insulin resistance. Novel effects connected adipose insubstantial biology. Mol Cell Endocrinol. 2021;536:111417.
Haase J, Weyer U, Immig K, Kloting N, Bluher M, Eilers J, et al. Local proliferation of macrophages successful adipose insubstantial during obesity-induced inflammation. Diabetologia. 2014;57:562.
Hill DA, Lim HW, Kim YH, Ho WY, Foong YH, Nelson VL, et al. Distinct macrophage populations nonstop inflammatory versus physiological changes successful adipose tissue. Proc Natl Acad Sci U S A. 2018;115:E5096–105.
Vandanmagsar B, Youm YH, Ravussin A, Galgani JE, Stadler K, Mynatt RL, et al. The NLRP3 inflammasome instigates obesity-induced inflammation and insulin resistance. Nat Med. 2011;17:179.
Kanda H, Tateya S, Tamori Y, Kotani K, Hiasa K, Kitazawa R, et al. MCP-1 contributes to macrophage infiltration into adipose tissue, insulin resistance, and hepatic steatosis successful obesity. J Clin Invest. 2006;116:1494.
Dommel S, Bluher M. Does C-C Motif Chemokine Ligand 2 (CCL2) Link Obesity to a Pro-Inflammatory State? Int J Mol Sci. 2021;22:1500.
Palmer AK, Xu M, Zhu Y, Pirtskhalava T, Weivoda MM, Hachfeld CM, et al. Targeting senescent cells alleviates obesity-induced metabolic dysfunction. Aging Cell. 2019;18:e12950.
Wang L, Wang B, Gasek NS, Zhou Y, Cohn RL, Martin DE, et al. Targeting p21(Cip1) highly expressing cells successful adipose insubstantial alleviates insulin absorption successful obesity. Cell Metab. 2022;34:75.
Matacchione G, Perugini J, Di ME, Sabbatinelli J, Prattichizzo F, Senzacqua M, et al. Senescent macrophages successful the quality adipose insubstantial arsenic a root of inflammaging. Geroscience. 2022;44:1941-60.
Sun K, Park J, Gupta OT, Holland WL, Auerbach P, Zhang N, et al. Endotrophin triggers adipose insubstantial fibrosis and metabolic dysfunction. Nat Commun. 2014;5:3485.
Marcelin G, Silveira ALM, Martins LB, Ferreira AV, Clement K. Deciphering the cellular interplays underlying obesity-induced adipose insubstantial fibrosis. J Clin Invest. 2019;129:4032.
Qi Y, Hui X. The shades of grey successful adipose insubstantial reprogramming. Biosci Rep. 2022;42:BSR20212358.
Winer DA, Winer S, Shen L, Wadia PP, Yantha J, Paltser G, et al. B cells beforehand insulin absorption done modulation of T cells and accumulation of pathogenic IgG antibodies. Nat Med. 2011;17:610.
Brestoff JR, Artis D. Immune regularisation of metabolic homeostasis successful wellness and disease. Cell. 2015;161:146.
Wensveen FM, Jelencic V, Valentic S, Sestan M, Wensveen TT, Theurich S, et al. NK cells nexus obesity-induced adipose accent to inflammation and insulin resistance. Nat Immunol. 2015;16:376.
McLaughlin T, Ackerman SE, Shen L, Engleman E. Role of innate and adaptive immunity successful obesity-associated metabolic disease. J Clin Invest. 2017;127:5.
Hildreth AD, Ma F, Wong YY, Sun R, Pellegrini M, O'Sullivan TE. Single-cell sequencing of quality achromatic adipose insubstantial identifies caller compartment states successful wellness and obesity. Nat Immunol. 2021;22:639.
Kratz M, Coats BR, Hisert KB, Hagman D, Mutskov V, Peris E, et al. Metabolic dysfunction drives a mechanistically chiseled proinflammatory phenotype successful adipose insubstantial macrophages. Cell Metab. 2014;20:614.
Snodgrass RG, Boss M, Zezina E, Weigert A, Dehne N, Fleming I, et al. Hypoxia Potentiates Palmitate-induced Pro-inflammatory Activation of Primary Human Macrophages. J Biol Chem. 2016;291:413.
Boutens L, Hooiveld GJ, Dhingra S, Cramer RA, Netea MG, Stienstra R. Unique metabolic activation of adipose insubstantial macrophages successful obesity promotes inflammatory responses. Diabetologia. 2018;61:942.
Lee YS, Olefsky J. Chronic insubstantial inflammation and metabolic disease. Genes Dev. 2021;35:307.
Lee BC, Kim MS, Pae M, Yamamoto Y, Eberle D, Shimada T, et al. Adipose Natural Killer Cells Regulate Adipose Tissue Macrophages to Promote Insulin Resistance successful Obesity. Cell Metab. 2016;23:685.
Li C, Menoret A, Farragher C, Ouyang Z, Bonin C, Holvoet P, et al. Single compartment transcriptomics based-MacSpectrum reveals caller macrophage activation signatures successful diseases. JCI. Insight. 2019;5:e126453.
Caslin HL, Bhanot M, Bolus WR, Hasty AH. Adipose insubstantial macrophages: Unique polarization and bioenergetics successful obesity. Immunol Rev. 2020;295:101.
Kintscher U, Hartge M, Hess K, Foryst-Ludwig A, Clemenz M, Wabitsch M, et al. T-lymphocyte infiltration successful visceral adipose tissue: a superior lawsuit successful adipose insubstantial inflammation and the improvement of obesity-mediated insulin resistance. Arterioscler Thromb Vasc Biol. 2008;28:1304.
Duffaut C, Galitzky J, Lafontan M, Bouloumie A. Unexpected trafficking of immune cells wrong the adipose insubstantial during the onset of obesity. Biochem Biophys Res Commun. 2009;384:482.
Nishimura S, Manabe I, Nagasaki M, Eto K, Yamashita H, Ohsugi M, et al. CD8+ effector T cells lend to macrophage recruitment and adipose insubstantial inflammation successful obesity. Nat Med. 2009;15:914.
O'Rourke RW, White AE, Metcalf MD, Winters BR, Diggs BS, Zhu X, et al. Systemic inflammation and insulin sensitivity successful obese IFN-gamma knockout mice. Metabolism. 2012;61:1152.
Ferno J, Strand K, Mellgren G, Stiglund N, Bjorkstrom NK. Natural Killer Cells arsenic Sensors of Adipose Tissue Stress. Trends Endocrinol Metab. 2020;31:3.
Toubal A, Kiaf B, Beaudoin L, Cagninacci L, Rhimi M, Fruchet B, et al. Mucosal-associated invariant T cells beforehand inflammation and intestinal dysbiosis starring to metabolic dysfunction during obesity. Nat Commun. 2020;11:3755.
Talukdar S, Oh DY, Bandyopadhyay G, Li D, Xu J, McNelis J, et al. Neutrophils mediate insulin absorption successful mice fed a high-fat fare done secreted elastase. Nat Med. 2012;18:1407.
Freitas DF, Colon DF, Silva RL, Santos EM, Guimaraes VHD, Ribeiro GHM, et al. Neutrophil extracellular traps (NETs) modulate inflammatory illustration successful obese humans and mice: adipose insubstantial relation connected NETs levels. Mol Biol Rep. 2022;49:3225.
Divoux A, Moutel S, Poitou C, Lacasa D, Veyrie N, Aissat A, et al. Mast cells successful quality adipose tissue: nexus with morbid obesity, inflammatory status, and diabetes. J Clin Endocrinol Metab. 2012;97:E1677–85.
Goldstein N, Kezerle Y, Gepner Y, Haim Y, Pecht T, Gazit R, et al. Higher Mast Cell Accumulation successful Human Adipose Tissues Defines Clinically Favorable Obesity Sub-Phenotypes. Cells. 2020;9:1508.
Khan T, Muise ES, Iyengar P, Wang ZV, Chandalia M, Abate N, et al. Metabolic dysregulation and adipose insubstantial fibrosis: relation of collagen VI. Mol Cell Biol. 2009;29:1575.
Datta R, Podolsky MJ, Atabai K. Fat fibrosis: person oregon foe? JCI Insight. 2018;3:e122289.
Grosse L, Wagner N, Emelyanov A, Molina C, Lacas-Gervais S, Wagner KD, et al. Defined p16(High) Senescent Cell Types Are Indispensable for Mouse Healthspan. Cell Metab. 2020;32:87.
Demaria M, Ohtani N, Youssef SA, Rodier F, Toussaint W, Mitchell JR, et al. An indispensable relation for senescent cells successful optimal coiled healing done secretion of PDGF-AA. Dev Cell. 2014;31:722.
Chaib S, Tchkonia T, Kirkland JL. Cellular senescence and senolytics: the way to the clinic. Nat Med. 2022;28:1556.
Fang J, Li L, Cao X, Yue H, Fu W, Chen Y, et al. Transmissible Endoplasmic Reticulum Stress Mediated by Extracellular Vesicles from Adipocyte Promoting the Senescence of Adipose-Derived Mesenchymal Stem Cells successful Hypertrophic Obesity. Oxid Med Cell Longev. 2022;2022:7175027.
Oh C, Koh D, Jeon HB, Kim KM. The Role of Extracellular Vesicles successful Senescence. Mol Cells. 2022;45:603.
Li Q, Hagberg CE, Silva CH, Lang S, Hyvonen MT, Salehzadeh F, et al. Obesity and hyperinsulinemia thrust adipocytes to activate a compartment rhythm programme and senesce. Nat Med. 2021;27:1941.
Sierra-Ramirez A, Lopez-Aceituno JL, Costa-Machado LF, Plaza A, Barradas M, Fernandez-Marcos PJ. Transient metabolic betterment successful obese mice treated with navitoclax oregon dasatinib/quercetin. Aging (Albany NY). 2020;12:11337.
Roden M, Shulman GI. The integrative biology of benignant 2 diabetes. Nature. 2019;576:51.
Nogueiras R, Sabio G. Brain JNK and metabolic disease. Diabetologia. 2021;64:265.
Regnier M, Van HM, Knauf C, Cani PD. Gut microbiome, endocrine power of gut obstruction relation and metabolic diseases. J Endocrinol. 2021;248:R67–82.
Gallego-Duran R, Montero-Vallejo R, Maya-Miles D, Lucena A, Martin F, Ampuero J, et al. Analysis of Common Pathways and Markers From Non-Alcoholic Fatty Liver Disease to Immune-Mediated Diseases. Front Immunol. 2021;12:667354.
Barreby E, Chen P, Aouadi M. Macrophage functional diverseness successful NAFLD - much than inflammation. Nat Rev Endocrinol. 2022;18:461-72.
Mashek DG. Hepatic lipid droplets: A balancing enactment betwixt vigor retention and metabolic dysfunction successful NAFLD. Mol Metab. 2021;50:101115.
Flessa CM, Kyrou I, Nasiri-Ansari N, Kaltsas G, Kassi E, Randeva HS. Endoplasmic reticulum accent successful nonalcoholic (metabolic associated) fatty liver illness (NAFLD/MAFLD). J Cell Biochem. 2022;123:1585-606.
Horn CL, Morales AL, Savard C, Farrell GC, Ioannou GN. Role of Cholesterol-Associated Steatohepatitis successful the Development of NASH. Hepatol Commun. 2022;6:12.
Huby T, Gautier EL. Immune cell-mediated features of non-alcoholic steatohepatitis. Nat Rev Immunol. 2022;22:429-43.
Ramadori P, Kam S, Heikenwalder M. T cells: Friends and foes successful NASH pathogenesis and hepatocarcinogenesis. Hepatology. 2022;75:1038.
Shaker ME. The publication of sterile inflammation to the fatty liver illness and the imaginable therapies. Biomed Pharmacother. 2022;148:112789.
Kolb H, Mandrup-Poulsen T. An immune root of benignant 2 diabetes? Diabetologia. 2005;48:1038.
Goldfine AB, Shoelson SE. Therapeutic approaches targeting inflammation for diabetes and associated cardiovascular risk. J Clin Invest. 2017;127:83.
da Cruz Nascimento SS, Carvalho de Queiroz JL, Fernandes de MA, de Franca Nunes AC, Piuvezam G, Lima Maciel BL, et al. Anti-inflammatory agents arsenic modulators of the inflammation successful adipose tissue: A systematic review. Plos one. 2022;17:e0273942.
Acknowledgements
I convey Stephan Martin, University of Düsseldorf, Germany, and Fraser W. Scott, the Ottawa Hospital Research Institute and the University of Ottawa, Canada, for reviewing the manuscript, and Kerstin Kempf, Düsseldorf Catholic Hospital Group, Germany, for helping with preparing the manuscript.
Funding
The enactment was supported by Gesellschaft von Freunden und Förderern der Heinrich-Heine-Universität Düsseldorf e.V.
Ethics declarations
Ethics support and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The writer declares nary competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with respect to jurisdictional claims successful published maps and organization affiliations.
Rights and permissions
Open Access This nonfiction is licensed nether a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, organisation and reproduction successful immoderate mean oregon format, arsenic agelong arsenic you springiness due recognition to the archetypal author(s) and the source, supply a nexus to the Creative Commons licence, and bespeak if changes were made. The images oregon different 3rd enactment worldly successful this nonfiction are included successful the article's Creative Commons licence, unless indicated different successful a recognition enactment to the material. If worldly is not included successful the article's Creative Commons licence and your intended usage is not permitted by statutory regularisation oregon exceeds the permitted use, you volition request to get support straight from the copyright holder. To presumption a transcript of this licence, sojourn http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the information made disposable successful this article, unless different stated successful a recognition enactment to the data.
About this article
Cite this article
Kolb, H. Obese visceral abdominous insubstantial inflammation: from protective to detrimental?. BMC Med 20, 494 (2022). https://doi.org/10.1186/s12916-022-02672-y
Received: 16 June 2022
Accepted: 21 November 2022
Published: 27 December 2022
DOI: https://doi.org/10.1186/s12916-022-02672-y
Keywords
- Obesity
- Adiposity
- Visceral fat
- Inflammation
- Adipocyte hypertrophy
- Adipocyte hyperplasia
- Adipose insubstantial macrophages
- Resident immune cells
- Cytokines
- Crown-like structures




 English (US)
English (US)